
In economia comportamentale parliamo di sconto intertemporale, avversione alle perdite, pesatura delle probabilità, reciprocità o punizione altruistica come se fossero parametri relativamente stabili della funzione di utilità.
Dal punto di vista biologico, però, questi parametri sono l’espressione comportamentale di sistemi neurali che si sono evoluti per risolvere tre problemi fondamentali: agire nel tempo, apprendere in condizioni di incertezza e coordinarsi con altri individui. Le preferenze non sono quindi oggetti primitivi: sono esiti emergenti di circuiti che integrano apprendimento, segnali corporei, controllo cognitivo e contesto sociale.
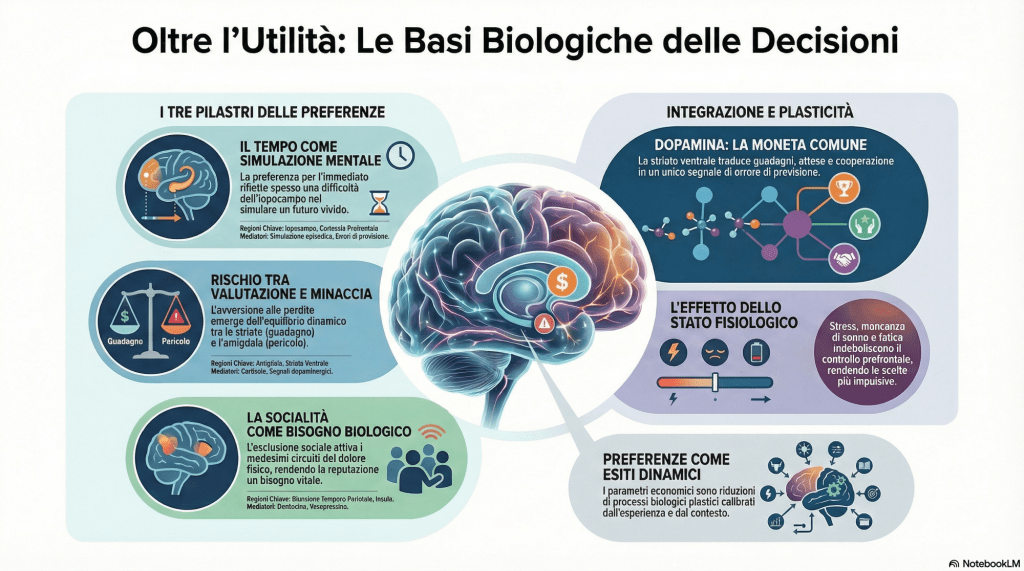
La rappresentazione del tempo è il primo pilastro. Il cervello non possiede un unico “orologio”, ma una rete distribuita che comprende gangli della base (in particolare lo striato), cervelletto e corteccia prefrontale. I neuroni dopaminergici del mesencefalo, noti per codificare gli errori di previsione della ricompensa, svolgono un ruolo cruciale anche nel collegare valore e tempo: segnalano quanto un esito differisca dalle aspettative e aggiornano le rappresentazioni del valore futuro. Ogni scelta intertemporale richiede due operazioni biologicamente costose: stimare l’affidabilità del futuro e simularlo mentalmente. Quest’ultima dipende in modo critico dall’interazione tra corteccia prefrontale e ippocampo, che consente la costruzione di scenari episodici dettagliati. Quando questa capacità è compromessa – per stress, privazione di sonno, fatica o immaturità dello sviluppo – il futuro diventa meno vivido, meno concreto, e quindi meno valorizzato. Ciò che osserviamo come “alto tasso di sconto” può riflettere una minore capacità di simulazione o un ambiente percepito come instabile, in cui gli errori di previsione sono frequenti e il sistema di apprendimento conclude che gli esiti futuri sono poco affidabili. In questo senso, la preferenza per l’immediato non è semplicemente impulsività: può essere la traduzione neurale di una credenza adattiva sulla volatilità dell’ambiente.
Il secondo pilastro è la gestione del rischio. Le decisioni rischiose attivano un’interazione tra sistemi di valutazione e sistemi di minaccia. Striato ventrale e corteccia orbitofrontale integrano magnitudine e frequenza degli esiti, apprendendo per esperienza attraverso segnali dopaminergici. L’amigdala, invece, è specializzata nella rapida individuazione di potenziali perdite o pericoli e può attivare risposte fisiologiche attraverso l’asse ipotalamo-ipofisi-surrene, con rilascio di cortisolo e aumento dell’arousal. Questa risposta aumenta la salienza delle perdite, contribuendo a spiegare perché, in molti contesti, le perdite “pesano” più dei guadagni. Non si tratta di un coefficiente universale inciso nel cervello, ma di un equilibrio dinamico tra reattività alla minaccia e capacità di regolazione prefrontale. La corteccia dorsolaterale supporta il controllo cognitivo e il calcolo deliberativo; la corteccia ventromediale integra segnali emotivi e corporei in un valore complessivo. In condizioni di stress o carico cognitivo elevato, il controllo si indebolisce e i segnali limbici dominano, producendo maggiore cautela o maggiore focalizzazione sul downside. Anche la pesatura non lineare delle probabilità può emergere da meccanismi di apprendimento per esperienza, campionamento limitato e memoria selettiva, senza che sia necessario postulare un calcolo esplicito delle probabilità: il cervello aggiorna aspettative in modo simile a un algoritmo di reinforcement learning, ma fortemente modulato dalla salienza emotiva degli esiti.
Il terzo pilastro riguarda la dimensione sociale. Valutare “gli altri” implica inferire intenzioni, aspettative e norme. Aree come la giunzione temporo-parietale e la corteccia prefrontale mediale sono cruciali per la mentalizzazione, ossia per attribuire stati mentali agli altri. Questa capacità è essenziale per la reciprocità condizionale: distinguere un atto ostile da un errore o da un vincolo esterno modifica radicalmente la risposta cooperativa. Le interazioni sociali sono inoltre regolate da sistemi neuroendocrini. L’ossitocina può aumentare fiducia e approccio sociale in contesti percepiti come sicuri, riducendo la reattività dell’amigdala a segnali minacciosi, ma tende anche a rafforzare il favoritismo verso il gruppo di appartenenza. La vasopressina è associata, in diversi contesti, a comportamenti difensivi o territoriali. Nei giochi di contrattazione, offerte percepite come ingiuste attivano l’insula anteriore, coinvolta nella rappresentazione di stati corporei avversivi, e la corteccia cingolata anteriore, che monitora il conflitto tra interesse personale e norma. Il rifiuto di un’offerta ingiusta, pur costoso, può quindi riflettere un valore intrinseco attribuito al rispetto delle norme e alla punizione di chi le viola. L’esclusione sociale, infine, attiva circuiti sovrapposti a quelli del dolore fisico, suggerendo che appartenenza e reputazione non siano semplici beni posizionali, ma bisogni profondamente radicati nella biologia.
Questi tre domini – tempo, rischio e sociale – convergono su circuiti comuni di valutazione, in particolare nello striato ventrale e nella corteccia prefrontale ventromediale, che integrano segnali temporali, probabilistici e sociali in un’unica rappresentazione di valore. La dopamina fornisce una “moneta comune” attraverso gli errori di previsione: un esito migliore o peggiore del previsto genera un segnale che aggiorna aspettative, indipendentemente dal fatto che si tratti di un guadagno monetario, di un risultato differito o di un’interazione cooperativa. Lo stato fisiologico – stress acuto o cronico, qualità del sonno, risorse energetiche – modula simultaneamente questi sistemi, alterando controllo, apprendimento e salienza della minaccia. Anche lo sviluppo è cruciale: i sistemi di ricompensa e sensibilità sociale maturano prima dei circuiti di controllo prefrontale, contribuendo a spiegare la maggiore impulsività “a caldo” e la forte influenza dei pari in adolescenza.
Da questa prospettiva, i parametri che stimiamo nei modelli economici sono riduzioni formali di processi biologici dinamici. Il tasso di sconto riflette la capacità di simulare il futuro e la credenza appresa sulla sua affidabilità; l’avversione alle perdite emerge dall’interazione tra sistemi di minaccia e regolazione; le preferenze sociali dipendono da meccanismi di mentalizzazione, segnali di sicurezza e norme interiorizzate. Le preferenze non sono dunque gusti immutabili, ma espressioni plastiche di sistemi neurali calibrati dall’esperienza e dallo stato dell’organismo. Comprendere questa base biologica non sostituisce i modelli economici, ma ne chiarisce la natura: ciò che appare come una funzione di utilità è, a un livello più profondo, il risultato di un cervello che integra tempo, incertezza e relazioni sociali per sopravvivere e coordinarsi in un ambiente complesso.
